
{kind=link}
Трансмембранная передача сигнала — свойство мембран — способность воспринимать и передавать внутрь клетки сигналы из внешней среды. Рецепторы:
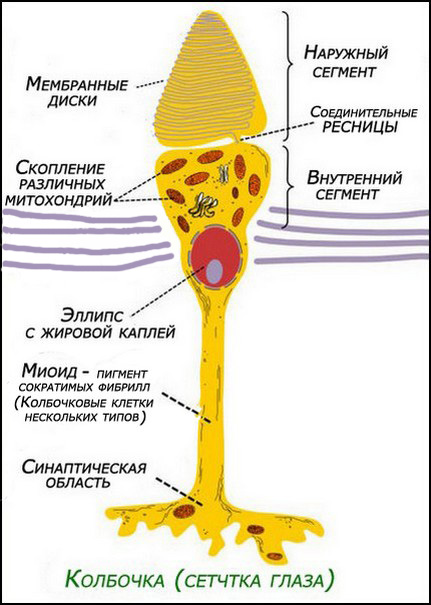
E=extracellular — Внеклеточное место — наружный сегмент мембраны колбочки, заполненный мембранными полудисками, образованными плазматической мембраной, и отделившимися от неё (обращённая к свету, наружная часть столбика из полудисков - постоянно обновляется, за счет фагоцитоза "засвеченных" полудисков клетками пигментного эпителия, и постоянного образования новых полудисков, в теле фоторецептора);
P=plasma (обработка и передача сигнала) — внутренняя поверхность рецептора;
I=intracellular — Внутриклеточное место — клеточная мембрана (см. рис.1,2).
Клеточный рецептор — молекула (обычно белок) на поверхности клетки, клеточных органелл или растворенная в цитоплазме, специфически реагирующая изменением своей пространственной конфигурации на присоединение к ней молекулы определенного химического вещества, передающего внешний регуляторный сигнал и, в свою очередь, передающая этот сигнал внутрь клетки или клеточной органеллы, нередко при помощи так называемых вторичных посредников или трансмембранных ионных токов.
Вещество, специфически соединяющееся с рецептором, называется лигандом этого рецептора. Внутри организма это обычно гормон или нейромедиатор либо их искусственные заменители, применяемые в качестве лекарственных средств и ядов (агонисты). Некоторые лиганды, напротив, блокируют рецепторы (антагонисты). Когда речь идет об органах чувств, лигандами являются вещества, воздействующие на рецепторы обоняния или вкуса. Кроме того зрительные рецепторы реагируют на свет, а в органах слуха и осязания рецепторы чувствительны к механическому давлению, вызываемому колебаниями воздуха и иными воздействиями.
Классы рецепторов[]
Клеточные рецепторы можно разделить на два основных класса - мембранные рецепторы и внутриклеточные рецепторы.
Мембранные рецепторы[]
Очень часто у белков есть гидрофобные участки, которые взаимодействуют с липидами, и гидрофильные участки, которые находятся на поверхности мембраны клетки, соприкасаясь с водным содержимым клетки. Большинство мембранных рецепторов - именно такие трансмембранные белки.
Многие из мембранных белков-рецепторов связаны с углеводными цепями, то есть представляют собой гликопротеиды. На их свободных поверхностях находятся олигосахаридные цепи (гликозильные группы), похожие на антенны. Такие цепочки, состоящие из нескольких моносахаридных остатков, имеют самые различные формы, что объясняется разнообразием связей между моносахаридными остатками и существованием α- и β-изомеров.
Функция “антенн” – это распознавание внешних сигналов. Распознающие участки двух соседних клеток могут обеспечивать сцепление клеток, связываясь друг с другом. Благодаря этому клетки ориентируются и создают ткани в процессе дифференцировки. Распознающие участки присутствуют и в некоторых молекулах, которые находятся в растворе, благодаря чему они избирательно поглощаются клетками, имеющими комплементарные распознающие участки (так, например, поглощаются ЛПНП с помощью рецепторов ЛПНП).
Два основных класса мембранных рецепторов — это метаботропные рецепторы и ионотропные рецепторы.
Ионотропные рецепторы представляют собой мембранные каналы, открываемые или закрываемые при связывании с лигандом. Возникающие при этом ионные токи вызывают изменения трансмембранной разности потенциалов и, вследствие этого, возбудимости клетки, а также меняют внутриклеточные концентрации ионов, что может вторично приводитъ к активации систем внутриклеточных посредников. Одним из наиболее полно изученных ионотропных рецепторов является н-холинорецептор.
{kind=link}
Структура G-белка, состоящего из трёх типов единиц (гетеротримерного) - αt/αi (голубые), β (красная) и γ (зелёная)
Метаботропные рецепторы связаны с системами внутриклеточных посредников. Изменения их конформации при связывании с лигандом приводит к запуску каскада биохимических реакций, и, в конечном счете, изменению функционального состояния клетки. Основные типы мембранных рецепторов:
- Рецепторы, связанные с гетеротримерными G-белками (например, рецептор вазопрессина).
- Рецепторы, обладающие внутренней тирозинкиназной активностью (например, рецептор инсулина или рецептор эпидермального фактора роста).
Рецепторы, связанные с G-белками, представляют собой трансмембранные белки, имеющие 7 трансмембранных доменов, внеклеточный N-конец и внутриклеточный C-конец. Сайт связывания с лигандом находится на внеклеточных петлях, домен связывания с G-белком — вблизи C-конца в цитоплазме.
Активация рецептора приводит к тому, что его α-субъединица диссоциирует от βγ-субъединичного комплекса и таким образом активируется. После этого она либо активирует, либо наоборот инактивирует фермент, продуцирующий вторичные посредники.
Рецепторы с тирозинкиназной активностью фосфорилируют последующие внутриклеточные белки, часто тоже являющиеся протеинкиназами, и таким образом передают сигнал внутрь клетки. По структуре это — трансмембранные белки с одним мембранным доменом. Как правило, гомодимеры, субъединицы которых связаны дисульфидными мостиками.
Внешние доли мембран фоторецепторов колбочек и палочек[]
{kind=link}
рис. 1. Колбочки (сетчатка)[1]

{kind=link}
рис. 2. Анатомия строения палочки (сетчатка)[2]

{kind=link}
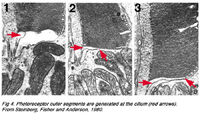
Рис. 4. Фоторецептор, внешние доли мембраны, созданные в реснице глаза (красные стрелки).[3]

{kind=link}
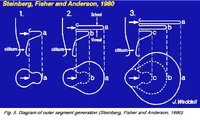
Рис. 5. Диаграмма внешней части мембраны с фоторецепторами[3]
Мембраны колбочек и палочек и их функция как биологические мембраны выполняют функцию восприятии пучков лучей предметной точки изображения c последующим оппонентным отбором основных лучей RGB в виде биосигналов (не в цвете, на рецепторном уровне) для передачи их в зрительные отделы головного мозга.
На основной части мембраны находятся места её выпячивания (evaginations), образующие внешнюю долю (o.s), наружный сегмент (см. рис. 1,2), которая является важной составляющей для обеспечения процессов визуализации, и содержит пигмент — как часть фоторецептора. Внешние доли палочек и колбочек являются результатом outpouching (a, рис. 5 ниже) мембраны — плазмы клетки фоторецептора в этом пункте (см. ниже) (Steinberg и др., 1980).
Эти расширяющиеся мембранные пластины (b-c, рис. 5 выше) в палочках выглядят как свободные, плавающие диски близ внешней мембраны. У колбочек внешние диски остаются сопряжёнными со внешней частью мембраны.[3].
Внутриклеточные рецепторы[]
Внутриклеточные рецепторы - как правило, факторы транскрипции (например, рецепторы глюкокортикоидов) или белки, взаимодействующие с факторами транскрипции. Большинство внутриклеточных рецепторов связываются с лигандами в цитоплазме, переходят в активное состояние, транспортируются вместе с лигандом в ядро клетки, там связываются с ДНК и либо индуцируют, либо подавляют экспрессию некоторого гена или группы генов.
Особым механизмом действия обладает оксид азота (NO). Проникая через мембрану, этот гормон связывается с растворимой (цитозольной) гуанилатциклазой, которая одновременно является и рецептором оксида азота, и ферментом, который синтезирует вторичный посредник - цГМФ.
Основные системы внутриклеточной передачи гормонального сигнала[]
Аденилатциклазная система[]
Центральной частью аденилатциклазной системы является фермент аденилатциклаза, который катализирует превращение АТФ в цАМФ. Этот фермент может либо стимулироваться Gs-белком (от английского stimulating), либо подавляться Gi-белком (от английского inhibiting). цАМФ после этого связывается с цАМФ-зависимой протеинкиназой, называемой так же протеинкиназа А, PKA. Это приводит к ее активации и последующему фосфорилированию белков-эффекторов, выполняющих какую-то физиологическую роль в клетке.
Фосфолипазно-кальциевая система[]
Gq-белки активируют фермент фосфолипазу С, которая расщепляет PIP2 (мембранный фосфоинозитол) на две молекулы: инозитол-3-фосфат (IP3) и диацилглицерид. Каждая из этих молекул является вторичным посредником. IP3 далее связывается со своими рецепторами на мембране эндоплазматического ретикулума, что приводит к освобождению кальция в цитоплазму и запуску многих клеточных реакций.
Гуанилатциклазная система[]
Центральной молекулой данной системы является гуанилатциклаза, которая катализирует превращение ГТФ в цГМФ. цГМФ модулирует активность ряда ферментов и ионных каналов. Существует несколько изоформ гуанилатциклазы. Одна из них активируется оксидом азота NO, другая непосредственно связана с рецептором предсердного натриуретического фактора.
цГМФ контролирует обмен воды и ионный транспорт в почках и кишечнике, а в сердечной мышце служит сигналом релаксации.
Фармакология рецепторов[]
Как правило, рецепторы способны связываться не только с основными эндогенными лигандами, но и с другими структурно сходными молекулами. Этот факт позволяет использовать экзогенные вещества, связывающиеся с рецепторами и меняющие их состояние, в качестве лекарств или ядов.
Так, например, рецепторы к эндорфинам — нейропептидам, играющим важную роль в модуляции боли и эмоционального состояния, связываются так же с наркотиками группы морфина. Рецептор может иметь, кроме основного участка, или «сайта» связывания со специфичным для этого рецептора гормоном или медиатором, также дополнительные аллостерические регуляторные участки, с которыми связываются другие химические вещества, модулирующие (изменяющие) реакцию рецептора на основной гормональный сигнал — усиливающие или ослабляющие ее, или заменяющие собой основной сигнал. Классическим примером такого рецептора с несколькими участками связывания для разных веществ является рецептор гамма-аминомасляной кислоты подтипа А (ГАМК). Он имеет кроме сайта связывания для самой ГАМК, также сайт связывания с бензодиазепинами («бензодиазепиновый сайт»), сайт связывания с барбитуратами («барбитуратный сайт»), сайт связывания с нейростероидами типа аллопрегненолона («стероидный сайт»).
Многие типы рецепторов могут распознавать одним и тем же участком связывания несколько разных химических веществ, и в зависимости от конкретного присоединившегося вещества находиться более чем в двух пространственных конфигурациях — не только «включено» (гормон на рецепторе) или «выключено» (на рецепторе нет гормона), а еще и в нескольких промежуточных.
Вещество, со 100% вероятностью вызывающее при связывании с рецептором переход рецептора в конфигурацию «100% включено», называется полным агонистом рецептора. Вещество, со 100% вероятностью вызывающее при связывании с рецептором переход его в конфигурацию «100% выключено», называется полным антагонистом рецептора. Вещество, вызывающее переход рецептора в одну из промежуточных конфигураций либо вызывающее изменение состояния рецептора не со 100% вероятностью (то есть часть рецепторов при связывании с этим веществом включится или выключится, а часть — нет), называется частичным агонистом или частичным антагонистом рецептора, в зависимости от преобладания агонистической или антагонистической активности. По отношению к таким веществам используется также термин агонист-антагонист. Вещество, не меняющее состояния рецептора при связывании и лишь пассивно препятствующее связыванию с рецептором гормона или медиатора, называется конкурентным антагонистом, или блокатором рецептора (антагонизм основан не на выключении рецептора, а на блокаде связывания с рецептором его естественного лиганда).
Как правило, если какое-то экзогенное вещество имеет рецепторы внутри организма, то в организме есть и эндогенные лиганды для данного рецептора. Так, например, эндогенными лигандами бензодиазепинового сайта ГАМК-рецептора, с которым связывается синтетический транквилизатор диазепам, также являются особые белки — эндозепины. Эндогенным лигандом каннабиноидных рецепторов, с которыми связываются алкалоиды конопли, является вещество анандамид, производимое организмом из арахидоновой жирной кислоты.
См. также[]
- G-белки
- Трансмембранные рецепторы
- Фоторецепторные клетки сетчатки глаза
- Рецептор (биология)
- Рецептор (биологическая клетка)
Примечания[]
- ↑ http://en.wikipedia.org/wiki/Cone_cell
- ↑ Human Physiology and Mechanisms of Disease by Arthur C. Guyton (1992) p.373
- ↑ 3,0 3,1 3,2 Helga Kolb. [http://webvision.med.utah.edu/book/part-ii-anatomy-and-physiology-of-the retina/photoreceptors/ Photoreceptors]. Webvision. Проверено 28 сентября, 2012.
| Это заготовка статьи по молекулярной биологии. Вы можете помочь проекту, исправив и дополнив её. |